
The Merriam-Webster Dictionary defines a time warp as “an anomaly, discontinuity, or submission held to occur in the progress of time.” How does this concept apply to life science research or the biotech industry? Two words succinctly summarize one pervasive time warp in life sciences: “lot testing.”
Lot-testing time is likely performed for the following reasons:
- Critical biochemical components responsible for the product’s biological effects are unidentified or unknown; or
- the key components causing the biological effect are identified, but the supplier does not have enough knowledge of the biochemical characteristics of the “factor(s)” to manufacture the components that generate the biological effect reproducibly;
- the supplier thinks they have control over manufacturing the factor(s), but a known or unidentified “contaminant” contributes to the biological effect.
The best illustration for lot selection is the supplementation of cell culture media with fetal bovine (FBS) or calf serum. The scientists developing defined culture media to grow mammalian cells in vitro in the 1950s rapidly realized that culture media required supplementation with ill-defined biological materials (e.g., serum, amniotic fluid) to enable cell growth.(1) FBS became the favored supplement because of its rich content of growth factors and hormones with low amounts of gamma globulin (i.e., antibodies).(2) This ill-defined material contains about 1800 proteins and more than 4000 metabolites.(3) One criterion for selecting a new lot is that the lot should generate a biological effect equal to or exceeding 80% efficacy of the current lot.(4) This criterion measures only one aspect of this complex biological material without knowing the components’ contributions. The development of serum-free media (5) to replace FBS was stimulated by the need to eliminate fetal bovine proteins that complicate the purification of therapeutic proteins generated during in vitro culture. Moreover, using defined media reduces the regulatory risk for the transmission of adventitious agents to patients.
Lot testing was also adopted in the 1960s to select “good” lots of Clostridium histolyticum collagenase for cell isolation. Research scientists found that collagenase could be used for tissue dissociation since it rapidly degraded nearly all forms of mammalian collagen, increasing cell yields from tissue. Worthington Biochemicals offered four Types of Collagenase (note, these have no relation to the mammalian collagen types). These products were manufactured by precipitating proteins in C. histolyticum culture supernatants with ammonium sulfate followed by resuspension of the protein pellet in a smaller buffer volume, creating crude collagenase enzyme preparations. These solutions were dialyzed to remove the salt and other low molecular weight contaminants. The dialyzed crude collagenase was characterized, bottled, lyophilized, and sold as Collagenase Types 1, 2, 3, or 4. Type 1 represented the most common collagenase and protease activities recovered after C. histolyticum fermentation. Types 2, 3, and 4 had higher protease, lower protease, and lower clostripain activities, respectively, compared to Type 1 collagenase.
Worthington, and later Sigma, provided small samples of specific lots of crude collagenase to cell isolators to determine if the new lot gave comparable results (e.g., cell yield, viability, function) to their present lot of collagenase. Analysis of the cell isolation results identified the lots as “good” or “bad” using the criteria that a new lot of collagenase gave comparable results to their current lot of collagenase used for cell isolation. Once a good lot of collagenase was determined, a larger amount of that specific lot was purchased, typically lasting > 1 year of cell isolations. Once the new lot was depleted, the lot qualification procedure was repeated.
Collagenase lot pre-qualification is understandable based on the biochemical heterogeneity of enzyme activities found in C. histolyticum culture supernatants. The primary biological effect of collagenase in tissue dissociation is to degrade the collagen in the extracellular matrix. Collagenases’ collagen degradation activity (CDA) loosens the extracellular matrix (ECM), exposing protease-sensitive sites on ECM proteins. Proteases in collagenase products cut the ECM proteins, including those “cell anchoring” proteins holding cells to the tissue.(6) Good lots contain the “right” amounts of collagenase and protease activities to release viable and functional cells from the tissue. The other enzymes appear to be superfluous contaminants.

Minimal control of the collagenase manufacturing process also contributes to the biochemical heterogeneity of crude collagenase products. Two different classes of C. histolyticum collagenase were identified after protein purification, each having a distinctive enzyme activity profile.(7) Class I (C1) collagenase had strong gelatinase and minimal peptidase activity when measuring the cleavage of a collagen-specific peptide substrate. In contrast, class II (C2) collagenase had weak to moderate gelatinase activity and strong peptidase activity.(8) Internal analysis of purified C1 collagenase at VitaCyte showed that C1 collagenase is more susceptible to proteolysis than C2 collagenase. C. histolyticum neutral protease and clostripain found in crude collagenase products can cut collagenase, affecting its ability to degrade collagen, as described below. In one report, Collagenase P (a crude collagenase) lost its ability to recover islets from porcine pancreata 18 months after product manufacture.(9) The loss of activity was likely caused by the high clostripain activity in this product since clostripain is very effective in cleaving C1 collagenase.
The development of the Liberase HI product for human islet isolation confirmed earlier observations that collagenase and proteases were the key enzymes responsible for tissue dissociation.(10) Specifically, HI contained ≈ 300 mg of class I (C1) collagenase, ≈ 200 mg of class II collagenase (C2), ≈ 58 mg of clostripain and ≈ 7 mg of thermolysin. This formulation was determined after analyzing a “great” lot of crude collagenase, Collagenase P, Lot 9. The purified pancreatic islets isolated using Lot 9 led to > 20 successful auto-islet human transplants performed at the University of Pittsburgh as part of a clinical research trial.(11) Thermolysin was used instead of C. histolyticum neutral protease, which was unstable after purification.
Liberase HI reduced the time leading islet transplantation centers spent to find good lots of collagenase. Several changes were deliberately made to the Liberase enzyme formulation during the product life cycle. In 2007, NIH discontinued using Liberase HI for any NIH-funded cell transplant clinical trials because of the potential risk of transmission of bovine spongiform encephalitis. NIH switched to Nordmark, a supplier that manufactured GMP-grade collagenase (Collagenase NB-1) for isolating human islets used in the Clinical Islet Transplantation Consortium clinical trial.(12) However, this change resurrected the “collagenase problem” because the most experienced islet transplantation centers participating in the NIH-funded clinical trials could not recover sufficient islets for transplant after they switched to the Nordmark enzyme.(6)
The discrepancy between the performance of the last version Liberase HI and Collagenase NB-1 for human islet isolation appeared to be due to the degradation of C. histolyticum C1 collagenase. (13) These analyses showed that collagenase purification alone was insufficient to maximize islet recovery from human pancreata. It is critical that C1 or C2 collagenase is protected from proteolysis during the manufacturing process. These comparative analyses concluded that collagenase-protease enzyme mixtures that contained intact C1 and C2 collagenase gave the highest islet yields. (13)
If your goal is to improve the consistency of your collagenase-mediated cell isolation process, the best place to break away from the collagenase time warp is to consider answering the three questions.
- Are you using intact C1 and C2 C. histolyticum collagenase? Does your collagenase supplier provide this information?
- Is the collagenase enzyme activity in excess to ensure effective collagen degradation in the tissue substrate? The best indication of insufficient collagenase CDA is large amounts of undigested tissue.
- Are you using a reasonable/optimal dose of neutral protease activity? The best indication of supraoptimal amounts of protease activity is damaged cells, low viability.
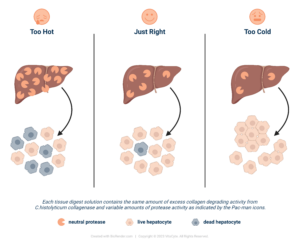
Successful enzyme-mediated cell isolations require an excess collagenase degradation activity in your tissue dissociation enzyme mixture. The use of collagenase that contains intact C1 and C2 ensures the consistency of the collagenase preparation and the likelihood that the CDA activity will be in excess. If these conditions are met, the neutral protease activity becomes a “regulator” to control the speed of digestion. If it is too hot, the cell will be damaged, and cell yield or function will be affected. Too cold, there is insufficient collagenase to loosen the ECM, which in turn does not expose the protease-sensitive sites on the cell anchoring protein that holds the cell to tissue. As Goldilocks would say, the protease needs to be just right to ensure the enzymes are not affecting your cell isolation process results.
If you have additional questions about these concepts, contact us.
If you have a specific application for development of purified defined enzyme mixture, contact Bob McCarthy at rcmccarthy@vitacyte.com.
References
1. Minonzio G, Linetsky E. The use of fetal bovine serum in cellular products for clinical applications: commentary. CellR4. 2014;2:e1307.
2. van der Valk J, Bieback K, Buta C, Cochrane B, Dirks WG, Fu J, et al. Fetal Bovine Serum (FBS): Past – Present – Future. Altex. 2018;35:99-118.
3. Chelladurai KS, Christyraj JDS, Rajagopalan K, Yesudhason BV, Venkatachalam S, Mohan M, et al. Alternative to FBS in animal cell culture-An overview and future perspective. Heliyon. 2021;7.
4. Even MS, Sandusky CB, Barnard ND. Serum-free hybridoma culture: ethical, scientific and safety considerations. Trends Biotechnol. 2006;24:105-8.
5. FCS free database F-F. 2023 [Available from: https://fcs-free.org/.]
6. McCarthy RC, Green ML, Dwulet FE. Evolution of enzyme requirements for human islet isolation. OBM Transplantation. 2018; 2: Available from: http://www.lidsen.com/journals/transplantation/transplantation-02-04-024.
7. Mandl I, MacLennan JD, Howes EL. Isolation and characterization of proteinase and collagenase from Cl. histolyticum. Journal of Clinical Investigation. 1953;32:1323-9.
8. Mookhtiar KA, Van Wart HE. Clostridium histolyticum collagenases: a new look at some old enzymes. Matrix Suppl. 1992;1:116-26.
9. Cavanagh TJ, Lakey JR, Wright MJ, Fetterhoff T, Wile K. Crude collagenase loses islet-isolating efficacy regardless of storage conditions. Transplantation Proceedings. 1997;29:1942-4.
10. Fetterhoff TJ, Cavanagh TJ, Wile KJ, Wright MJ, Dwulet FE, Gill J, et al. Human pancreatic dissociation using a purified enzyme blend. Transplantation Proceedings. 1995;27:3282-3.
11. Ricordi C, Tzakis AG, Carroll PB, Zeng YJ, Rilo HL, Alejandro R, et al. Human islet isolation and allotransplantation in 22 consecutive cases. Transplantation. 1992;53:407-14.
12. Clinical Islet Transplantation Consortium 2018 [Available from: http://www.citisletstudy.org/.
13. Balamurugan AN, Breite AG, Anazawa T, Loganathan G, Wilhelm JJ, Papas KK, et al. Successful human islet isolation and transplantation indicating the importance of class 1 collagenase and collagen degradation activity assay. Transplantation. 2010;89:954-61.